预计阅读本页时间:-

第8章 可塑的感官
精妙又机敏的大脑啊,你比我更聪明。
——威廉·卡洛斯·威廉斯
思想家很早就知道感觉是“可塑”的。你常常能看到,如果剥夺一个人的一种感觉,其他感觉就会有所代偿,例如盲人的听觉和触觉会增强。此外,知觉学习实验(我们将在第9章中讨论这类实验)表明,一个人的感觉的敏锐度可以通过练习获得很大的改善。但是,持怀疑态度的人可能会说,这仅仅是注意力、专注度和练习的问题,而不是真正的感官改善。我们必须使用现代方法来证明大脑神经元的回路的确发生了物理变化。
广告:个人专属 VPN,独立 IP,无限流量,多机房切换,还可以屏蔽广告和恶意软件,每月最低仅 5 美元
受损的感觉会重塑
“大脑可塑性”这个术语指大脑重组其环路的能力。早期的经典实验是这样的:你有一只天生没有视力的大鼠(举例来说,它一出生你就损毁了它的视网膜),等它长大后,你训练它跑迷宫。在它学会跑迷宫之后,你再轻微损坏它的视觉皮质,然后让它再次跑迷宫。(大鼠很乐意跑迷宫,因为大鼠能在迷宫的末端得到食物奖励。)你比较了大鼠在视觉皮质损坏前后的跑动速度。原则上,对一只已经失明的大鼠来说,损坏它的视觉皮质似乎不会产生任何影响,但是你却发现大鼠走迷宫的表现确实变差了,这表明盲大鼠的视觉皮质对迷宫奔跑有所贡献,尽管我们不知道具体是什么贡献。该实验由耶克斯灵长类动物生物学实验室的卡尔·拉什利(Karl Lashley)等人完成,他以寻找记忆的物理位置而出名。之后的研究者指出了拉什利研究的局限性,但事实证明,他的工作方向是正确的。[1]
同一时期,临床医生在人类患者身上有了新的发现,这次是两类发育诱发的失明。第一类患者从出生起一只眼睛就被闭塞了——由于白内障或罕见的眼睑问题,后来,虽然解剖上的问题解决了,但这只眼仍然瞎了或近乎瞎掉。早年的闭眼经历使那只眼睛无法正确连接到其中枢神经通路。
第二类发育诱发的失明,发生在两只眼睛天生指向不同方向的孩子身上。这些孩子长大后,你经常发现他们的一只眼睛接管了另一只眼睛:一只眼睛在工作,另一只眼睛却没有,这就是弱视。另一只眼睛并不是真正瞎了——你可以通过专门的测试证明它的视网膜仍在起作用,但没有有用的视觉通过这只眼睛进入大脑。(目前有多种针对这种情况的疗法,最常见的方法是在幼年时用眼罩交替遮挡两只眼睛,以使一只眼睛永远没有机会接管和压制另一只眼。)
被称为视觉皮质图像处理发现者的先驱戴维·休伯尔和托斯坦·维泽尔决定在动物中重复这些实验,并发现了弱视的神经基础:在一段关键的年龄时期,从视网膜输出到中枢神经系统的突触是可塑的。如果皮质神经元从一只眼睛得到大量信号,而另一只眼睛却没有,那么第一只眼的轴突会吸引所有皮质神经元的注意力(即它们的突触连接),从而使得第二只眼即使功能正常,也没有皮质神经元可以与之对话。
他们发现,对于斜视患者来说,情况则有些微妙。在正常情况下,一只眼睛的图像和另一只眼睛的图像几乎完全重合,并且视觉场景中的一个点会刺激一组皮质神经元。休伯尔和维泽尔人为地让动物的眼睛交叉指向不同方向,例如让幼年动物戴上会改变其视觉图像的棱镜,使得两只眼睛的图像无法正确地聚焦在同一目标上。如果两只眼睛的视线偏离,以至于不能完全平行,那么视野中央的视觉输入就会偏移,不再能形成单个皮质映射图。这个人看到了字面意义上的重影——看到两个单独的图像。在眼睛交叉的情况下,大脑面临一个问题:两只眼睛的图像相互冲突,它必须选择一只眼睛或另一只眼睛。于是不被选择的那只眼睛的连接便受到了抑制。这种抑制起初是暂时的,但过了一段时间之后,那只眼睛就永久性地处于失明状态了。
一项巧妙的实验证明了另一种发生在皮质上的重组。在正常情况下,视觉皮质上有视网膜的“映射图”。虽然皮质表面的起伏肯定会扭曲这张图,但是你还是能一眼看出视网膜上相邻的点也会投射到视觉皮质上相邻的点上,从而在其上创建视觉场景的组织图。实验者使用激光在猴子的视网膜上无痛地打一个非常小的洞。然后,洛克菲勒大学的查尔斯·吉尔伯特(Charles Gilbert)记录下了视觉皮质的反应。最初,皮质图上也有一个对应于视网膜上洞的空白,但是,过了一会儿,邻近区域的皮质反应迁移并占据了空缺,也就是说视网膜临近受损区域的细胞开始与皮质当中原本对应受损区域的皮质神经元沟通了。
这并不意味着视网膜受损区域恢复了视力。如果你的视网膜有病变,那么你永远不会通过受损区域看到任何东西——你有了一个盲点。但是,即使大脑永远无法弥补视网膜上的孔,视网膜病变周围的区域也会比以前“拥有”更多的皮质神经元,这预示它将做得更好。据我所知,这一预测尚未得到检验。不过至少,受更多神经掌控的区域应该会更坚挺,可能会比以前更耐损伤。
理解这一现象的角度之一,是自然界会防止皮质闲置。假设皮质某一区域不再从其自然位置接收输入,那么该皮质区域永远不活动将会造成浪费。取而代之的是,过一会儿,它的功能就会被交给未损坏的输入。在一般情况下,你可以轻松地将这一机制想象为一种处理小中风的方式。(神经病理学家告诉我们,在我们的生活中,所有人都会遭遇这些小的脑组织损失。)想象一下,你的皮质只是小中风,只影响很小的血管,而它所喂养的大脑区域却死去了。对于过去从该区域中受损的区域接收输入的大脑区域来说,这将是永远的沉默,这将浪费宝贵的皮质资源。取而代之的是,大脑会将那些大脑区域分配给其邻居,以充分利用糟糕的情况。
重塑正常知觉
在上一节中,我们看到了感官是如何适应各种类型的神经损伤的,这些损伤对于神经系统来说规模巨大且粗暴。但还有些自然发生的微妙重组,会发生在我们所有人身上。
实验人员能从脑部扫描中学到很多。在天生失明的人的大脑活动中,我们能看到显著的大脑可塑性。当盲人志愿者在扫描仪中用手指阅读盲文时,通常处理视觉输入的大脑区域(如初级视觉皮质)被激活。这些人多年来一直在密集地使用触摸。不知怎的,触觉信息的处理已经占据了未使用的视觉中心。
还有一个戏剧性的例子,来自视觉正常的小提琴家。要拉小提琴,你需要用一条手臂大幅度、粗略地运动,让弓上下扫过琴弦;另一只手则进行一系列非常细微的动作,在小提琴的指板上下飞舞,快且多变地按压琴弦。如果你是一位出色的小提琴家,这些动作会很快,如果你是顶级演奏家,那按弦速度会更快。这项任务所需的速度和精度是很了不起的,专业的小提琴家每天练习这些动作数小时之久。这对他们大脑中的物理连接产生了影响。你可能已经猜到了,这是因为手指的运动受特定大脑区域的控制。对专业的小提琴家来说,其相关脑区会不断扩大,甚至使邻近脑组织的功能丧失。但这仅发生在用于按弦的手上。控制另一只手的大脑另一侧的相同区域没有扩展,因为即使在专业演奏者身上,那只手需要做的动作也相对粗糙。手臂大幅运动那一边(即拉琴弦的手臂那一边)控制手指运动的大脑相关区域完全保持正常。
(小提琴家是一个极端的例子,但我想知道在其他情况下会发生什么。如果你是一名职业运动员,你控制肌肉的大脑回路会以牺牲别的回路为代价来扩张吗?如果你的大部分工作时间都在担心大脑,那么担心大脑的回路会以牺牲欣赏歌剧的回路为代价扩张吗?)
在实验室中可以测试相反的情况,即感官剥夺。在黑暗中长大的猫不能正确地融合两只眼睛的图像。还有一个尚存争议的实验,实验组里的幼猫只被允许看垂直条纹或水平条纹的图案。动物出生时神经元的方位选择性很弱,随着动物的成长,神经元的方位选择性会变得更敏锐。在只能看到条纹的极端情况下,长大后动物的初级视觉皮质神经元的方向选择性会偏移:异常多的细胞会偏好特定的朝向——如果猫唯一的视觉体验是垂直条纹,这个特定朝向就是垂直的,如果猫只看到水平条纹,则为水平的。[2]
这类实验还有一个巧妙的变种,是在动物幼时剥夺其观察物体运动的能力。实验人员用闪光灯间歇短暂地照亮环境,将猫置于其中饲养。这使猫可以看到平常的世界,但是灯光闪烁的时间太短,无法使物体在视网膜上发生任何有意义的运动,这样一来,这些动物的视觉皮质也体验不到任何物体的运动。这样做会发生什么呢?这些动物长大后,其皮质中没有方向选择性神经元。
最终和关键的一组实验操作直接证实了突触可塑性在视觉发育中的作用。这些实验分别由迈克尔·斯特赖克(Michael Stryker)、卡拉·沙茨(Carla Shatz)和他们的学生完成,这些学生现在分别在加州大学旧金山分校和斯坦福研究LGN。
LGN的一部分专用于从一只眼睛输入,另一部分专用于从另一只眼睛输入。但是,我们出生时LGN并非以这种方式开始。在正常婴儿中,来自两只眼睛的轴突广泛分布,每个都覆盖了LGN的大部分。一开始左右眼的输入几乎没有分离。分离是由从眼睛来的轴突的活动模式产生的。甚至在婴儿出生之前,这些轴突就在发放——就像汽车发动机怠速运转时那样,以簇状形式突突地发放[3]。这些突突的发放是同步的:来自同一只眼睛的活动总是同时到达LGN,而另一只眼睛的活动则在另外的时间到达。
这为一种关键的可塑性形式(它也启发了机器学习)创造了条件。正如你将在第9章中了解到的,唐纳德·赫布提出,同时发放的神经元群之间的联系会增强。当一只眼睛的许多视网膜轴突同时驱动其LGN目标细胞时,相对于另一只眼睛的突触,这些轴突和LGN细胞之间的突触会增强。渐渐地,最初胡乱分布的轴突精细化了它们的LGN目标,于是,一团LGN神经元对来自右眼的输入做出了响应,而另一组对左眼的输入做出了响应。结果,LGN的左右眼映射图变得清晰分明,这靠的正是增强同时活跃的突触。为了证实这一点,斯特赖克使用药物阻止一只眼睛发出的活动到达LGN,在这种情况下,LGN上两眼的输入就不再分开。
所有这些发现都表明感觉系统的可塑性。但是,在自然条件下,这有多重要?如果一个人在没有视力的情况下成长,会发生什么?
学着去看
赫布认为视觉主要是后天习得的:复杂的感知都是通过体验的联结形成的,因为世界上的物体,其视觉特征都是成群出现,而不是一个一个孤立的。他认为这些联结必须发生在生命的早期,否则,正如当时的某些证据所表明的那样,之后大脑将无法形成必要的细胞集群。他的基本想法是正确的:视觉的形成的确依赖于体验。但是他关于这必须在年轻时发生的结论似乎仅是部分正确的。
证据来自那些天生失明、后来又通过治疗重获视力的人。麻省理工学院的帕万·辛哈(Pawan Sinha)是印度人,在一次回国访问时他意识到,在印度乡村中,可能有30万名儿童患有先天性白内障。在这些儿童中,眼睛里的晶状体被浑浊的纤维组织所代替。白内障的眼睛能穿透光明与黑暗,但这些儿童丧失了更精细的视力。在人道主义与科学的完美结合下,辛哈组织了一个慈善项目,寻找这些孩子并将他们送到新德里,在那里,现代医院的外科医生用透明的人工合成的晶状体替换了他们的晶状体。在发达国家,医生给许多患有白内障的老年人实施了相同的手术。
辛哈的团队在手术前、术后,以及数月或数年后检查了患者的视力。摘除白内障并不能立即恢复儿童的精细视力。世界在他们眼前混乱而模糊。但是随着时间的流逝,他们的视力开始改善。几个月后,他们能看到更多细节。许多人可以不用手杖走路了,能在拥挤的街道上骑自行车、认识朋友和家人、上学,并且开展各种需要视力的活动。
然而,他们的视力似乎从未变得完美。即使经过几个月的训练,他们的视力仍低于正常水平。有人评论说他可以看报纸上的头条新闻,但看不到小字号的正文。有些人在执行特定的视觉任务时遇到了麻烦,例如,将彼此重叠的两种形状分开,如下图所示。
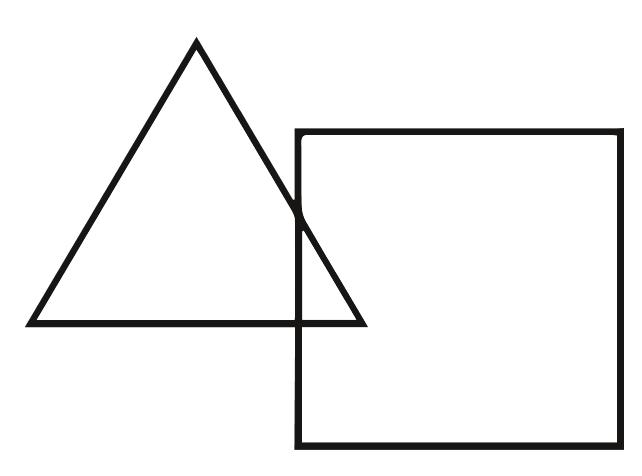
我们大多数人都将这张图片看作与正方形部分重叠的三角形,但是一些新近获得视力的人只将这些线条组成的图案视为单个复杂的对象。(有趣的是,如果你让三角形或正方形彼此独立地移动,那么此问题就会“得以解决”。这种形式的视觉训练和其他形式的视觉训练似乎有助于视觉的恢复。)
这么看来,大部分视力都可以在长大后恢复。但是有一些发现提醒我们,视觉系统的可塑性不是无限的。首先,脸部识别斑块在不同的人(或猴子)中位置相似这一事实表明,它们在大脑中的出现具有某种程度的内在规律。其次,随着手术后的印度儿童逐渐恢复视力,他们的大脑活动模式发生了变化。白内障摘除后,功能性磁共振成像显示,他们的皮质对各种视觉输入(包括面部)的反应混乱而分布广泛,但很快,对面部有反应的区域就只剩下一系列斑块,这些斑块正位于它们该在的位置。这再次表明,大脑提前知道了面部补丁应该在哪里。至少在一定水平上,这是视觉结构由先天决定的证据。利文斯通将这些预先决定的位置称为“原型脸部斑块”。
最后,利文斯通和她的同事于2017年年底发表了一项关于感觉神经可塑性的优雅而有力的实验。他们饲养的猴子从小就没见过面孔,不仅仅是没见过人的面孔或猴子的面孔,应该说从来没有见过任何面孔。完成这项实验听起来可能需要做很多工作,但原则上并不太难。实验人员亲切地照顾了小猴子,无论何时,只要他们靠近它们,实验人员都会戴上电焊工的面罩——这是一块很大的深色玻璃的弯曲板,从额头伸到下巴下方。
除了看不到脸,这些猴子在一个完全正常的视觉世界中长大:它们可以看到笼子和周围房间中的一切;它们可以看到实验人员的躯干、手臂和脚;它们可以看到喂奶的奶瓶。它们可以听到猴群发出的正常声音。它们唯一被剥夺的是对面部的视觉体验。这些猴子在大多数情况下都能正常发育,并且在实验完成后被引入猴群时,它们能与同伴开心地交流,而且成功地融入了猴子社会。
实验人员训练了这些猴子,使其能安静地躺在fMRI扫描仪内。之后,他们向猴子展示了包括面孔在内的各种视觉物体,并对其大脑进行了扫描。你可能已经猜到了,它们脑中并没有面部斑块。不过,值得注意的是,通常原本是颞叶面部识别脑区的区域会对手的图像做出反应。在正常的社交环境中,灵长类动物最重要的视觉对象是面孔。面孔表达愤怒、恐惧、敌对、爱以及所有对生存和繁荣至关重要的情感信息。显然,环境中第二重要的特征是手——猴子自己的手,以及培育和喂养它们的实验人员的手。
尽管通常会成为面部斑块的脑区变成了“手部斑块”,但它对视觉物体的偏好仍然具有可塑性。在猴子被允许看到实验人员和其他猴子的脸后大约6个月,脸部斑块中的细胞逐渐恢复为对脸敏感。显然,面部传达的信息如此重要,以至于它们重新夺回了手的图像所占据的大脑区域。
面部斑块的存在解释了一个早就被认识到的奇怪临床症状——字面意义上的“脸盲症”(源于希腊语“脸”和“无知”)。这些人的视力很正常,但是难以识别人脸。患者可以和其他人一样区分一张脸和另一张脸,但是却很难从记忆中识别出脸。
脸盲症的程度不同——有非常轻微的,也有极端严重到需要去看医生的。而另一个极端是,有些人是超级人脸识别器。据身边的一位助手说,爱德华·肯尼迪参议员可以认出一万人。就个人而言,我其实更接近脸盲一端。这是一个令人尴尬的问题。我可能前一天与你共进晚餐,度过一个愉快的夜晚,第二天在大厅见到你时却想:“我认识那个人吗?”没有什么办法确认。因此,如果我一次又一次怠慢你,请你理解,这是我的问题,而不是我对你没有兴趣。
总结一下:面部斑块在颞叶广泛分布,似乎可以协同工作,并通过体验变得对面部敏感。正如利文斯通所指出的,这个分布广泛、依赖于经验的系统,在很多方面都表现得很像是一个可以习得的神经环路。
[1] 卡尔·拉什利(1890—1958)是神经科学的先驱,他深入思考了大脑结构和行为的关系。他曾经做了一系列实验来定位大脑区域和记忆之间的关联[所谓大脑里的记忆痕迹,Semon把它称为“印迹”(engram)]。拉什利是赫布的研究生导师,也是他一生的朋友。他的研究工作是赫布理论的根基。拉什利的工作被他总结在自己的经典论文中:Lashley,K.S.(1950).In search of the engram.In Society for Experimental Biology (Ed.),Physiological mechanisms in animal behavior (Society’s Symposium IV) (pp.454–482).Oxford,UK:Academic Press.
[2] 20世纪70年代时,围绕这些实验的争议沸沸扬扬。争论基本上是聚焦在两派之间,即以休伯尔和维泽尔为代表的天生派和唐纳德·赫布为代表的环境塑造派。天生派认为感受野背后的神经连接是出生前就编程好的,而环境塑造派则认为感受野受到视觉刺激的强烈影响。休伯尔和维泽尔发表文章报道了猴子的视觉皮质自出生就有方位选择性,这反驳了赫布的观点,不支持线条检测是神经元学习的结果。我见过休伯尔寄给赫布的一封唐突——我并不是说他粗鲁——的信件,里面用完全确定的词汇描述了这个结论。之后又有一项研究表明V1的一些细胞在猕猴非常年幼,还来不及接受很多视觉输入时就有方位选择性了,还有研究表明从小没有见过任何轮廓的动物也能发展出方位选择性。这使得赫布的看法显得不可信。然而,尽管休伯尔在很多问题上都做出了正确的判断,但在这个问题上却可能错了。很快,科学家了解到,新生动物的脑内只有少数细胞有方位选择性,这些细胞的数量在物种间存在差异(一些动物出生时比另一些动物拥有更多方位选择细胞),而且这些细胞的方位选择性也比成年动物脑中细胞的方位选择性要低。参见Espinosa,J.S.,& Stryker,M.P.(2012).Development and plasticity of the primary visual cortex.Neuron,75,230-249.
[3] 如果你考过C1或其他手动挡机动车驾照,或是观察过起步时的燃油公交车,你应该会熟悉那种突突突地颤抖的感觉。——译者注










