预计阅读本页时间:-

第4章 幽灵神经元
忒瑞西阿斯,如果你明白,你就得门儿清,否则你就是不明白。[1]
——埃兹拉·庞德
21世纪的神经科学有一场悄无声息的革命,它重塑了解剖学研究。解剖学一度被认为是无聊的、无用的操作,是“蝴蝶标本采集”。然而,了解大脑的结构一直都很重要。神经科学的开创者和守护神——圣地亚哥·拉蒙–卡哈尔(Santiago Ramón y Cajal)的研究就完全基于神经解剖学。可以肯定的是,医学生最不愿意做的事情之一,就是去背诵大脑的几十条神经束和几十个神经核团。然而宽泛地说,神经解剖学——或者按照现在的说法,有时叫作结构神经生物学——正是神经科学的核心所在:大脑是一个连接而成的机器,大脑做的所有事最终都归结到它的各个部分互相连接的方式。
广告:个人专属 VPN,独立 IP,无限流量,多机房切换,还可以屏蔽广告和恶意软件,每月最低仅 5 美元
2000年左右,几项技术进步叠加在一起,为理解大脑解剖结构带来了巨大的突破。第一项进步是显微镜分辨率的巨幅提升,标志是共聚焦显微镜的发明。另一项进步是让细胞组分可视化的方法爆炸性地增多了。分子生物学的神奇工具让我们能为最小的亚细胞组分制造标记,而共聚焦显微镜让我们能看到它们:在自然环境中游动着的运动细胞;在黑暗下成团的发光细胞,不同细胞类型还能发不同的光[2]。这些进步让我们能做以前不敢想的梦:普查大脑中的所有神经元——列出大脑(或视网膜)的完整“组成表”,这是解开连接奥秘的第一步。
无法鉴定的神经元
海因兹·瓦塞尔在佛罗里达海边会议上所做的报告,让我们这些神经生物学家知道,可以自下而上地将视网膜拆解开来研究。我们可以制作一个完整组成表,然后探究每个组分在干什么。就像我刚刚说的,我们很快有了很棒的新工具来帮助我们。
免疫组织化学(immunocytochemistry)在1990年左右出现,它是一种能让我们看到几乎所有蛋白质分子在细胞或者组织内位置的工具。你在电视上看到闪闪发亮的旋转着的神经元,大多数是用免疫组织化学看到的,这个技术操作上较简单,呈现出的结果在视觉上又很华丽。
它也可以让人挫败。我可以告诉你一个故事,一款商用试剂浪费了我实验室一年的工作(从物质的角度,可以说一家不道德的供应商让美国纳税者白花了30万美元)。然而,神经生物学家还是一头扎了进去。朱莉·桑德尔(Julie Sandell)一开始在哈佛,后来去了波士顿大学;哈维·卡滕(Harvey Karten)和尼克·布雷查(Nick Brecha)本身就是技术先驱;瑞典的伯恩特·埃因格尔;德国的海因兹·瓦塞尔和利奥·佩奇;得克萨斯的戴安娜·雷德本(Dianna Redburn)和史蒂夫·马西(Steve Massey)……新西兰的戴维·瓦尼(David Vaney)用显微镜拍下了美丽的图片,找到了他原初的爱:他在变老以前退出了科研圈,做起了职业摄影师。
只要有合适的试剂,任何人都能用一台荧光显微镜照亮所有包含特定分子的视网膜神经元。在较低的放大倍数下,你能在黑暗背景中看到一片闪亮的星星。在较高倍数下,你能看到一个神经元的具体形态,它那细细的突起穿进视网膜中,这些突起是神经元之间互相连接之所在。哪些试剂会挑出哪些亚型的视网膜神经元里的分子呢?这通常是(现在仍然是)靠猜测的。如今最好的探针还是突触的神经递质:多巴胺、我们的老朋友乙酰胆碱、血清素等,它们中的每一个都在相对较少种视网膜神经元中存在。(当然,一个神经元里包含的分子种类更多,或许有上万种,但它们中的大多数在许许多多种神经元都存在,不仅仅是视网膜,在大脑和身体别处也有;它们做一些例如提供能量、维持结构之类的事。对我们的研究来说,它们用处不大。)
我们实验室发表了二三十篇研究论文,收集了一条视网膜神经元类型的列表(大约有12种)。这些细胞类型的染色很可靠,你可以轻易重复这些实验。这意味着这些类型的所有细胞,都可以与视网膜里的其他神经元在显微镜下区分开。我们可以测量它们的大小,可以给它们计数——这听上去或许微不足道,但却是真正的科学的基础。这让我们能超越标本采集,超脱于单一“典型”细胞的概念,转向一幅这些细胞关于视觉做了什么、又不做什么的图景。举个例子,一些类型的视网膜神经元数量很少,但却散布在视网膜各处。我们知道这样的细胞不会去传输高分辨率的图像:低细胞数量意味着很少的像素,细胞数量太少,每个细胞采样范围太大,以至于其传输给大脑的图像清晰度将大大降低,每个点都是巨大的模糊泡泡。相应地,有些小细胞数量很多,紧密地堆积在视网膜上。我们不难想到它们将从视网膜向大脑提供一条高分辨率通路,事实也的确如此。
我们和其他实验室美滋滋地拍摄着精美的图片,猜测视网膜的每个部分如何工作。然而过了不久,我们就没有分子可染了。只有一些分子能标记特定类型的神经元,我们试过的其他分子都不行。不幸的是,房间里还有头大象:我们能鉴定的大多数细胞数量相对较少,我们可以标记整群整群的细胞类型,而被染上颜色的那些细胞都散布在整个视网膜上,彼此隔得很远。在它们之间的空隙里,有整排整排的神经元,我们用过的任何分子都不能把它们标记染色。如果说视网膜是一本涂色本,我们只能给20%的部分涂上颜色,剩下的全是空白。
这令人不安。对我们所尝试的研究来说,我们要理解神经节细胞的所有信息源,这是至关重要的:如果大部分潜在的提供信息的细胞都不在我们的视线中,我们怎么能指望理解视网膜的计算呢?怎么去理解对比增强、方向选择以及其他种种呢?
我得承认,我们试图将这些细胞分类出来的最大动机还是出于好奇心。假如给你一台古董钟表让你维修,却不给你操作手册,也许你用不了多久就能看出钟摆是计时结构的一部分,但是面对躺在里面的一堆黄铜齿轮、转盘,你怎么才能知道它们各自派上了什么用场呢?大自然这位超凡的钟表匠,就是这么捉弄我们的。
面对视网膜或是中枢神经系统的其他部分,问题就在于没有特殊染料的时候,所有神经元看上去都一样。我们的通用染料只能标识细胞体,但是正是那没有染上的细细突起——伸出去接收输入的树突,以及向其他神经元发送信号的轴突,才让每种细胞类型变得特殊。因此,对神经细胞类型的研究才一度是标本采集般的逸事记录,解剖学家只能研究刚好被染上的个例,而一个细胞能否被染上靠的完全是运气和直觉。
我们觉得在视网膜上我们应该能做得更好,因为它在结构上有起始有终结,信息流动方向又是单一的。与许多大脑区域不同的是,我们知道视网膜的功能是什么,而且它在空间上也很紧凑——从光感受器到神经节细胞只有约0.3毫米。我们觉得,为视网膜里的所有细胞绘制一张地图,而不仅仅是随机寻找标本,应该是一个可以实现的目标。用现代术语来说,这种列出一整个结构里所有神经元的研究会被称作神经组学(neurome),这是在类比基因组学(将一个动物的所有基因列出来的研究)。
追踪幽灵神经元
但该怎么做呢?我们先从最基础的层级开始。惊人的是,当时我们只有视网膜神经的最粗糙分类:光感受器、水平细胞(horizontal cell)、双极细胞、无长突细胞以及神经节细胞。用普通的染色方法,这五种细胞看上去差不多,与本页下方插图中的这些卵圆形小圈没什么差别。我们知道这些细胞大类的存在,也可以猜测它们大致的数量比例,但是怎么才能搞清楚那边实际有什么呢?下图展现了视网膜在我们眼前的大致样子:我们能鉴定少数细胞(涂黑的、有细长突起的细胞),但剩下的空心圆圈,对我们来说是个谜。
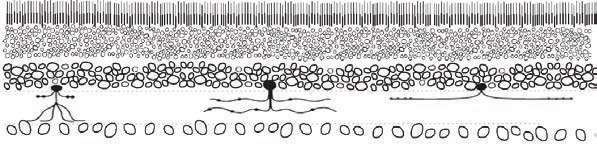
为了寻求建议,我找到了埃利奥·拉维奥拉(Elio Raviola),他是哈佛神经生物学系的资深成员。拉维奥拉是科学家中的科学家,精通神经解剖学中的一切。我问,他的十八般武器之一——电子显微镜(electron microscopy),是否能用来鉴定不同类型细胞之间的区别。他说当然能,但是这很费人工,得有个人坐在切片机前切几万张视网膜的超薄切片。埃利奥还有更棒的事情要做,帮不了忙,但他向我推荐了意大利科学家恩里卡·斯特雷托伊(Enrica Strettoi),她刚在他那里做完一轮博士后。和埃利奥一样,恩里卡是一个聪明、友善、外向,而且在科学上毫不含糊的人。他们一起完成过一个超棒的、用连续切片电子显微镜研究视网膜内神经连接的项目。恩里卡为我的项目带来了技术、规范和激情,她的关键性洞见让我们的研究得以成功。
“我们并不需要用高分辨率分析来折磨自己,”她说,“与其纠结于细微的差别,不如用根本的定义来鉴定细胞,也就是它们的突起向哪个方向延伸、在哪一层形成连接。”在电子显微镜下,这些突起很大。事实上,恩里卡指出,把光学显微镜的分辨率推到极致的话,我们也能在镜头下看到它们。这样的话,我们就能少切很多切片,因为光学显微镜的切片厚度可以比电子显微镜所要求的厚10倍,而它覆盖的视野要大得多。于是,我们准备了三维的视网膜固体样本(这在电子成像时代很容易,在当时则难以想象),并且切成一系列切片。我们的目标是把样本里每一个细胞都确定无疑地鉴定出来。
我们在波士顿准备视网膜样本。回到比萨后,恩里卡把它们切成连续切片,给每张切片拍照。随后,她将底片邮寄给我们。那时,邮寄靠的还是国际平邮(这个项目几近完成时,我们总算用上了数字底片和电子邮件)。我们小组的第三位成员是我的技术员丽贝卡·罗克希尔(Rebecca Rockhill)。丽贝卡是我们的劳模,我让她去暗房冲洗几千张相片,她二话不说,只花了5个星期就从暗房出来了,手里拿着的8.5英寸乘11英寸的相片还散发着显影剂的呛鼻烟雾。
我们坐在一张长桌上把那堆相片一张一张地翻过去,来鉴定每一个细胞。如果你和我们坐在一起,你会看到,在第一张照片上,有一堆细胞体在不同位置切出的不规则侧面。你选一个细胞,随便选一个,然后翻到第二张相片找同一个细胞,看到它在稍微不同的深度被切出的侧面。你再翻到第三张,重复这个操作,直到你看到一根突起离开细胞体——一根树突或轴突。你问你自己:“这个突起会向上延伸到光感受器细胞,还是向下延伸到神经节细胞?”通过下一张相片,你更加明确了突起的方向。你继续翻,看着突起延伸到内突触层或外突触层,再过几张切片,这根突起便缩小不见了。这时你已经从细胞体一路找到这根突起的起始和终点,直到它太细,细到你再也找不下去了,但你已经跟了它足够远,你的自信足以判断出它到底延伸至视网膜内层还是外层。
毋庸置疑,鉴定细胞一根突起的轨迹,意味着你能通过细胞类型的根本定义鉴定它是双极细胞、无长突细胞还是水平细胞(见第77页图):无长突细胞只把突起延伸到视网膜内层,水平细胞只延伸到外层,而双极细胞则会延伸到两层。那让我们回到第一张相片,用标签笔在细胞体上写上“B”(双极细胞)、“A”(无长突细胞)或“H”(水平细胞)。如果这是一个细胞类型中我们第一个标记的细胞,那我们就会写上“B1”、“A1”或“H1”,然后再看下一个细胞。
我自己鉴定了一些细胞,其他的则由暑期实习的学生完成。(这听上去像是一个无聊的暑期实习,但学生们似乎并没有受到什么永久性创伤,其中有两位如今还是功成名就的神经科学家。)因为我们对鉴定的每个细胞都在纸上做了标记,所以我们确保自己能回过头检查我们的结论,这样,每一步都是严谨的,我们把视网膜中层的所有细胞都鉴定了个遍,明确了水平细胞、双极细胞和无长突细胞的确切占比。这个实验没有出差错的空间,这感觉很棒。
一旦我们明确了细胞的大类,我们就可以问一个更细节的问题:我们之前的染色漏掉了多少无长突细胞?我们先从无长突细胞开始,是因为它们是视网膜内层中数量最多却被研究得最少的细胞。在所有无长突细胞中,有多少细胞是我们已知的类型?令人震惊的是,所有已知的类型加起来只占细胞总数量的24%。[3]
尽管有一个清楚的答案很好,但这个答案并不让人振奋。还记得我们最初的目标吗,为什么我们要数细胞数到深夜?我们是要理解视网膜如何处理信息:视网膜通过神经节细胞给大脑发送了什么信息,也就是说,视网膜是如何产生视觉的第一步的?如果我们想知道神经节细胞是如何产生它的输出的,那对我们来说,看不见神经节细胞剩下的76%潜在输入,真是太不走运了。
恩里卡·斯特雷托伊
意大利国家研究委员会神经科学研究所的恩里卡·斯特雷托伊,身高约1米6,穿着得体,笑口常开。即使她有时穿牛仔裤来实验室,身上也总会有些创意穿搭——她从不穿T恤,而且她常常穿着一双高跟鞋。在我的记忆里,恩里卡从仲夏比萨的拥挤街头走来,镇定自若,穿着优雅的白色亚麻夹克和点缀着珍珠的短裙,脚上的高跟鞋轻松地踩过并不平整的石子路。
她生长在比萨,现在还住在那里。比萨是中世纪欧洲最早的大学城之一。她在母亲的杂货铺的楼上长大。如今,她住到了郊区的大房子里,房屋外是围墙花园,这是一幢她丈夫家所有的农场墅院。丈夫卢卡是她邻居家的孩子。她的母亲和另外两个女儿住在附近,其中一个女儿是医生,另一个是仍在受训的医学生。与她们热情洋溢的母亲不同,女儿们安静,说话轻声细语,讲起英语时流利又小心。
恩里卡工作很努力。像意大利人推崇的那样,她热爱在周末为家人烹饪;其他时候则比任何人来实验室都早,走得却很晚。她努力对蠢人礼貌,但不会太有耐心。她的习惯是激情工作几个月,然后和家人过一个长长的假期——8月在海滩或阿尔卑斯山,圣诞节在比萨的家里。她是一个虔诚的天主教徒。她和卢卡都在当地的歌剧团唱歌。她给朋友发邮件时会在结尾附上“来自恩里卡的拥抱”。
她的科研目标之一是缓解几种常见的遗传性盲症,这些疾病被统称为色素性视网膜炎(RP),病因是视网膜光感受器细胞里的基因缺陷。如果你遗传了其中一种基因,你的光感受器细胞就会退化,你就会变瞎——有时是在出生后几年,有时是几十年后。
恩里卡想知道感觉输入是否会影响退化进程。她和学生把拥有同样基因缺陷的小鼠作为模式系统来研究。她把一组小鼠养在无聊的普通笼子里,另一组几乎同样的小鼠养在她说的“充满玩具”的笼子里:有可供攀爬的木头格子,可供躲藏的洞,可供运动的跑轮。让她吃惊的是,在充分丰容的环境里,小鼠的视网膜确实退化得慢得多。长话短说,事实证明,主要的益处来源于那个跑轮——运动,同时结合了感觉刺激。[4]
这具体是怎么做到的尚不清楚,学界对恩里卡的发现也没有给予太多的关注。运动有益健康并非新闻——大家都知道运动是“神药”,可以延缓甚至预防从头到脚的各种疾病。但是对于任何罹患RP的患者来说,运动能延缓视网膜退化这件事值得为他们所知,我可不喜欢他们不知道。因为恩里卡的科学总是做得那么完美无缺,我相信她的数据最终会被更多人所了解。如果我因为视网膜退化而开始变瞎,我保证每天会在跑步机上跑两次。
围堵无长突细胞
我们怎么才能抓住这76%鬼影般的无长突细胞呢?翻阅免疫化学标记的目录已经不能让我们找到什么新东西了。我们需要一些鉴定视网膜细胞的无偏方法,让我们能看到所有的细胞类型。
解决方案分成三部分。第一步,我们决定不用分子标记,转而观察细胞的形状。神经元树突和轴突的精致分叉形状自神经科学的黎明时期起就牢牢吸引了科学家。确实,卡哈尔笔下优雅的神经元树形结构,近期成了麻省理工学院一场艺术展览的主题。一直有些人怀疑神经元的形状也许不代表什么意义——它也许反映了细胞的发育历史,但不代表它成熟后的功能。然而,神经细胞的形状有一个无可辩驳的重要性:它反映了神经元的突触连接。
在下页图中,可以从侧面看到三种视网膜神经元(A、B和C),就像你切开眼球看到的剖面边缘。细胞A和细胞C是无长突细胞(根据定义,它们只有延伸到内视网膜的短突起,内视网膜是更靠近神经节细胞的那面)。注意,它们的突起延伸到内视网膜的不同层。这至关重要。无长突细胞C不可能与神经节细胞B有突触连接,因为它们的分叉分布在不同层。只有相互接触才能形成突触连接,这两种细胞显然接触不到彼此。
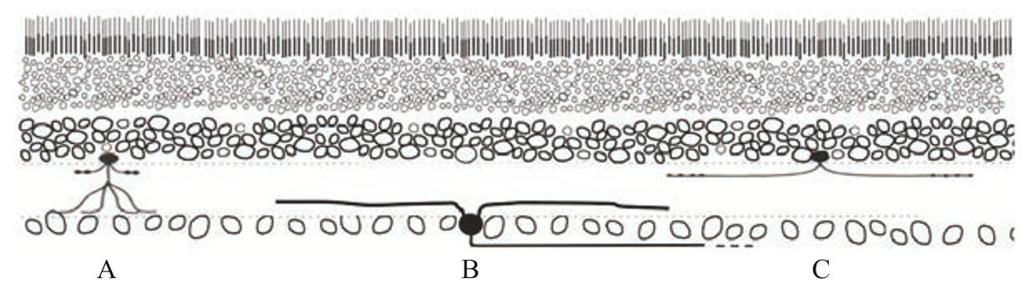
第二件值得注意的事情是,细胞突起伸展得很广,无长突细胞A和C肯定执行的是不同的功能,因为它们的形状不同,所以它们一定是两种类型的细胞。细胞A很小,细胞C很大。一个视网膜细胞的侧向伸展范围决定了它看的视野有多大。有些细胞树突伸展得广,有些伸展得窄,前者看到世界的更大一片,后者看到的是一小片。无长突细胞A和C执行着不同的视觉任务,向视网膜神经节细胞发送着不同的信号,也就让神经节细胞发送给大脑的信息变得不同。
但首先,我们怎么看到细胞的形状呢?上图所展示的细胞整体形状,在一般情况下是看不见的。细胞体——它是细胞的大本营、DNA(脱氧核糖核酸)的储藏室,也存放着让细胞产能和建造其他细胞结构的机器——还是很容易看到的,但是细胞的突起——树突和轴突,不那么容易看到,它们很细,而且更重要的是它们和其他细胞的突起纠缠在一起。即使你有一个能染所有树突的染料,你也不能确定它们来自哪些细胞。
我们需要一个能让单个细胞从其他细胞中脱颖而出的手段。不仅如此,这个方法还得可控,让我们能用它来系统性地从所有无长突细胞中采样。我们最终选定的技术是光填充(photofilling),这需要把视网膜泡在一种光敏分子溶液中,让这些分子渗透进每个细胞。然后,我们用一束集中的光,照射出一个比细胞还小的光斑,打在随机选定的一个无长突细胞上。在这个细胞内部,这一个小亮斑将触发一连串反应,让荧光分子在整个细胞里扩散,从而令这个细胞从数百万个没有荧光的背景细胞中脱颖而出。
这是一个麻烦的方法。举个例子,我们不能用常规手段拍摄荧光图像了,因为我们用来激发荧光的激发光会让周围的细胞也发生反应,发出荧光。我们只好买了一台极端敏感(也更昂贵)的数码相机,让我们能在不到0.1秒内拍照,周围细胞的荧光分子来不及在快门关闭前扩散。这个方法在小细胞上表现得比大细胞更好,不过通过练习,我们手艺高超的博士后玛格丽特·麦克尼尔(Margaret MacNeil)能够达到非常高的成功率。事实上,她随便选一个细胞打光,94%的时候都能拍摄到树突丛的照片。这意味着随机取样几百次就能得到所有类型无长突细胞的代表性画像了。
还记得我们这个研究的原初问题吗?如果我们能鉴定的24%无长突细胞实际上是稀有类型,那常见的无长突细胞是什么类型呢?让我们很吃惊的是,答案是根本没有常见类型的无长突细胞。
这意味着什么呢?我们曾经希望找到无长突细胞中的一些主要角色,其他无长突细胞则扮演着周围的配角。然而出乎我们所料的是,无长突细胞是一群多样的细胞,且各种类型分布均匀。这样的话我们只能假设它们在处理视觉信息方面具有同等的重要性,我们的论文发表在一本重点期刊上(发表时只遇到了一点儿难度),结论是视网膜里有29种不同的无长突细胞,它们在处理视觉图像上各自发挥了不同的作用。
为什么这个微小的结论如此重要?事实证明,这是揭示视网膜如何工作的重要线索。视网膜究竟为什么需要29种无长突细胞?答案一定是因为视网膜中存在着比我们想的多得多的信息处理。无长突细胞是视网膜神经节细胞的主要输入之一,而神经节细胞是视网膜向大脑发送信息前的最后一步。如果无长突细胞如此多样,神经节细胞发送的信息一定也相当多样。这是一个进步,在理解视觉上的一个小的进步。
幽灵细胞续:双极细胞
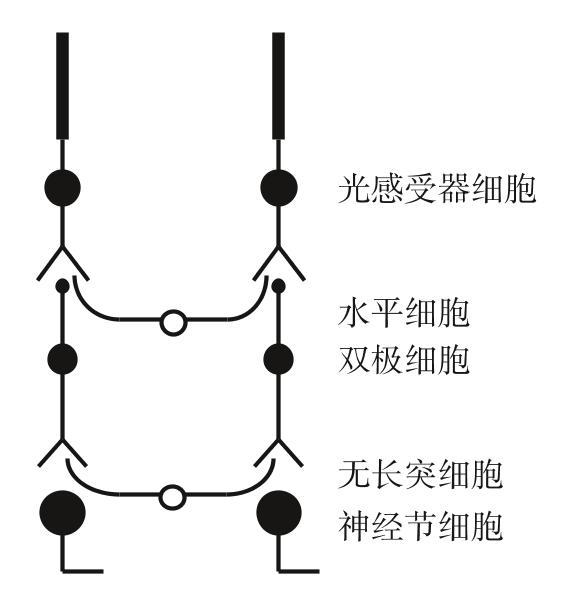
我们研究无长突细胞的同时,另一些伙计也在摆弄着视网膜里的微环路。那时最重要的知识空白是双极细胞。如果你记得的话,双极细胞从光感受器细胞处接收突触输入,向视网膜里伸出一根突起,与无长突细胞和神经节细胞形成突触连接。它们是视网膜里的重要成员。如果你从视网膜里取出所有的无长突细胞,视网膜仍然能干活,虽然有点儿勉强:瞬态和持久神经节细胞依然会存在,不过它们之间的差别将不再明显;也不再会有方向选择性细胞了。一个没有无长突细胞的人还可以看,虽然他的视觉将会变得模糊且迟钝。但如果你把所有双极细胞都拿掉,视网膜就只能分辨白天黑夜了,这还得靠一小部分本身就对光敏感的神经节细胞。
每个人都听说过科学突破——新发现或新想法像划破天空的一道闪电一样袭来,改变一整个科学领域。这绝不是常有的事。科学进步靠的是积淀:证据的积累让可能变成可信,让可信变成事实。这也是在我们理解视网膜双极细胞的道路上发生的事。
对双极细胞的首次系统记录,来自金子章道(Akimichi Kaneko)、弗兰克·韦布林(Frank Werblin)和约翰·道林(John Dowling)。记录显示双极细胞似乎有4种类型:瞬态开,瞬态关,持久开,持久关。很自然就能想到,这4种双极细胞驱动了4种神经节细胞。
但是,和无长突细胞类似,我们也有理由相信,双极细胞不止已被发现的这4种类型。到20世纪90年代中期,有四五个实验室开始研究双极细胞,他们对细胞类型的估计从4到9不等。我自己的实验室很晚才赴宴。我们意识到前人的工作给了我们一个粗略的概念,让我们知道视网膜双极细胞大概是怎么组织的。但是这些经验很大程度上都基于逸事型的记录,也就是之前批评的“蝴蝶标本采集”,这个类型记下几个例子,再换一个类型记下几个。于是我们问了一组不同的问题:第一,有没有之前的技术染不上的双极细胞?第二,有没有什么细胞类型占主导地位?是不是有一些“老板”细胞,被一些“帮手”细胞围绕?还是说所有的双极细胞的贡献都差不多?
为了回答这些问题,我们找来了埃利奥·拉维奥拉。他对视网膜做过一组精美的染色,但是那些切片当时已经在他实验室束之高阁了好一阵。[5]他的一个学生对它们做了一些初步研究,但是这个项目还是被抛弃了,因为埃利奥这样的完美主义者敏锐地知道,不管他的手艺多么稳定,很有可能还是有双极细胞没有被染上。
我的实验室为这个项目带去了两样东西:我们贡献了光填充技术,这个技术让我们能无偏、无变地将所有双极细胞都纳入镜中。我们还给了他们玛格丽特·麦克尼尔,她当时是神经元三维成像大师。她做出的图是我们的快乐源泉,解剖学家热爱神经元的漂亮图片。我们从神经元的照片里感受到了神秘,我们觉得展现在我们面前的,就是真理的一部分。
我们甚至还有第三样指导我们分类的信息:当时细胞电突触已经被发现,有一种分子能标记出电突触之所在,这样我们就可以先标记双极细胞的电突触再成像。这部分工作是我们的朋友,阿拉巴马大学的雷·达舍(Ray Dacheux)所做的。
知道细胞对光的实际反应是无价的补充,因为细胞的反应和它们的形状一样截然分明。三种方法(染色、光填充和微注射)的偏差各有不同,这样的话,很难想象有什么细胞能在三种方法下都逃脱。结合所有信息,我们对自己的结果有了更多信心。我们认为,双极细胞共有13种,它们长得如下图,为埃利奥所画:

这幅图片强调了双极细胞的鉴定特征——它们轴突延伸的深度。我们的发现也被他人证实,即双极细胞间最大的不同就是它们的轴突在视网膜的哪个突触层分叉。如果你还记得之前那张无长突细胞的示意图,不同深度的内视网膜突触层有不同的其他细胞——无长突细胞和神经节细胞——的树突,它决定了一个双极细胞与哪种无长突细胞和神经节细胞接触。
我们发现,没有一种双极细胞占主导。就像无长突细胞一样,所有类型的细胞都或多或少地均等分布。也就是说,从光感受器到内视网膜,有大概13条平行的通路,然后它们会遇到大概29种无长突细胞,以及更多数量的神经节细胞,从而生成给大脑的视觉输入的最终密码。
随着时间推移,人们发明了更好的染色方法。海因兹·瓦塞尔和他的学生用免疫染色技术完成了另一件杰作,对双极细胞进行了更新的普查。他们的计数更加精确仔细,他们能把每种细胞的数量都数出来,加起来和双极细胞总数(我和恩里卡的计数结果)吻合。瓦塞尔和他的同僚总结道:“我们的目录完整收录了11种视锥双极细胞和1种视杆双极细胞,小鼠视网膜的所有主要双极细胞类型似乎都已经被发现了。”[6]尽管如今有了更先进的电子显微技术和更强大的遗传分子标记,我们也只对麦克尼尔和瓦塞尔实验室做的目录做了微小修改。取决于不同的标准,细胞类型总数被定在12到15种不等。
这十几种双极细胞是视网膜的核心,它们代表了“初级”视觉。视网膜和之后的脑区域将这些双极细胞送出的信息以不同组合拼凑、修改,强调其中一部分,忽略另一部分。但是大脑不能逾越双极细胞所设定的上限。双极细胞就是视觉的基本信息源。
布赖恩·博伊克特
20世纪末,视网膜领域最有影响力的学者是布赖恩·博伊克特,他是一名英国王家学会会士,却从未被授予过任何高等学位。
我第一次遇到他是在他位于伦敦德鲁里巷的国王学院的实验室。[7]那是一个积灰的、看起来空荡荡的地方。木制的书柜,合页已经不灵活了,里面摆着古老的笔记本。布赖恩那时已是一位著名的科学家,他是生物物理研究院院长,因被埋没而闻名的女科学家罗莎琳·富兰克林(Rosalind Franklin)正是在这个学院拍下了著名的DNA结构的X射线衍射图,成就了沃森和克里克。布赖恩穿着朴素的衬衫和陈旧的宽松长裤,没戴领带。他的肚腩垂到了皮带以下,嘴里抽着没有滤嘴的香烟。我当时是个紧张而野心勃勃的入门者,刚刚开始为校外的同行所知。我和他面对面坐在实验室的椅子上。没有磨蹭多久,布赖恩就问起了我的实验,他已从小道消息对此有所耳闻。我们的谈话持续了好长一段时间,从此以后,我们还有更多次烟味缭绕的漫长对话。
1924年的冬天,布赖恩·博伊克特在英国克罗伊登出生。他7岁时,他的母亲带他离开了酗酒的父亲,这让母子俩在大衰退时期失去了经济支柱。他们和朋友住了几个月,随后他的母亲找到了一个薪水不高的工作,得以租下一个单间。
幸运的是,布赖恩的生父曾是共济会成员,这让小男孩得以被共济会的一所教会学校接收。那是一所典型的英国寄宿制学校,为他提供了食物、庇护甚至衣服。他在8岁入学,除了短暂的假期,他的童年基本上在共济会学校度过。
布赖恩的童年还被疾病困扰。在学校里,虽然他似乎并没有过得不快乐,但却学什么都不在行。他法语和数学不及格,好不容易才过了物理和化学。他被剑桥大学拒绝,只能去伯贝克学院进修。伯贝克学院是伦敦大学的一个分支,是为了“在夜间教育工匠阶层”而创立的,换句话说,这是一所夜校。那时正值“二战”,是伦敦大轰炸的高峰。伯贝克学院被炸,授课只能在建筑的残骸的地下室中进行,楼上满是皱卷的废铁。伦敦大学的日间学生已经被疏散到了北威尔士的舒适安全地带,但是博伊克特和他的夜班同学只能在锡皮屋顶下学习,这里夏天热得像烤箱,下雨时则吵得让人耳聋。
布赖恩四处寻找日间工作,最终在动物学系的动物房做上了助手,他的第一个任务是打扫笼舍。也许是否愿意做这份脏活是一个测试,因为他很快就被调到了一个更有趣的职位上。他成了生理学实验室的一名低阶技术员。
在前一年,这个生理学实验室还是亨利·戴尔爵士(Sir Henry Dale)领导的。戴尔是突触生物学的先驱,他退休后,实验室保留了原本的结构和学科。布赖恩四年的本科生涯,白天扑在实验室,晚上留给上课。这对一个成长期的生物学家来说是非凡的经历。他在动物房结识了英国的工人阶级,他尊敬并喜爱他们。在戴尔的实验室,他结识了科学精英。那个年代的研究组的规模还比较小,即使是世界闻名的戴尔,也只雇用了不到15个同事和技术员。我想这个年轻能干的小技术员一定特别招人喜欢,因为他很快就有了自己的实验可做。其中之一是把狗放在平台上摇晃,直到它们呕吐。这个研究是王家空军要求的,显然是为了理解晕机背后的基础生物学原理。布赖恩没有告诉我晕机实验本身的结果,但他告诉我,有一只聪明的狗学会了一看到实验装置就吐,好让自己不再参与当天的实验。布赖恩之后持续研究巴甫洛夫条件反射背后的神经生物学原理,兴许也不是巧合了。
后来布赖恩在书中感谢了这段生活经历,那时他自己搭建并使用实验装置,结识了志同道合的科学家。他在这一时期写出了自己的第一篇科学论文,描述了一种测量蛙人呼吸装置里二氧化碳积累量的新方法。不幸的是,军方将此研究列为“机密”,尽管当时战争已经结束,博伊克特的第一篇研究成果还是没能发表。
但是很显然,他给自己的老板留下了深刻的印象。从伯贝克毕业后,布赖恩在伦敦大学学院谋得了动物学助理讲师的职位。在那里,他的主要任务是负责教授导论课程的实验课。这个基础而微小的工作如今都交给研究生助教做了。他注册了博士项目,但是没有就读,因为很快他就得到了在世界著名动物学家约翰·扎里哈·杨(J.Z.Young)处做研究助理的机会,工作地点是杨在意大利那不勒斯的海洋生物实验室。博伊克特认为杨的指导是唯一能让他学习动物如何学习的机会。他要研究的是章鱼的大脑,大海一直以来都为动物学家提供了一大堆可供研究基础原理的简单动物。
他们对于学习的神经基础的研究广受好评,不仅在大众媒体上被广泛报道(章鱼的学习,引人注目),也被各个科学领域所知。但是唯一的坏处是,相比博士论文,布赖恩的名人老板更希望他写出下一篇科学论文。直到最后布赖恩也没拿到博士学位。不过同时,年轻的博伊克特获得了很高的科学声誉。在这种情况下,英国的学术权威破格地认为博士学位对布赖恩来说并不重要,布赖恩由此得到了教职。之后,他总是很快地纠正别人不要叫他博伊克特博士,并自豪地表示自己唯一正确的称谓是博伊克特教授。
从那不勒斯回到英国后,他开始对其他与学习有关的课题感兴趣,其中之一是松鼠冬眠时大脑发生了什么变化。他搬到哈佛做了半年教学工作,部分原因是为了收集松鼠,波士顿的地松鼠可比伦敦多得多。
他在哈佛遇到了约翰·道林,他们之间的合作推动了科学进步。道林刚开始做一个用电子显微镜研究脊椎动物视网膜的项目,但是电子显微镜的放大倍数太高了,道林虽然看到了视网膜内迷人的突触排布,却不能在这么大的放大倍数下找到伸出这些突触的细胞。博伊克特是全细胞染色专家,在他研究章鱼的过程中,他就已经很擅长解开神经环路的迷宫了。博伊克特和道林很快意识到了他们之间的互补性。他们一起发表了一篇有关视网膜连接的标志性研究论文,为之后的解读铺平了道路。
布赖恩能看穿大图景,如激光般瞄准下一个大问题。在视网膜这个领域,他只带着一名技术员做了一个有关视网膜精细结构的研究。但他对这一领域的最大贡献不是他亲自做的研究,而是给我们这些后辈带来了看待学科的视野的广度。布赖恩每个月在瓦塞尔的法兰克福研究所做顾问,做批评家,做编辑,做教父以及拉拉队。瓦塞尔是一位优秀的科学家、一位高效的管理者,而且有马普所的实验室资源供他调配。因此当布赖恩思考时,瓦塞尔的实验室就会努力完善实验的细节。尽管不是一代人,博伊克特和瓦塞尔两个人还是很相似,他们既有活力又有纪律性,既正直又勤勉。
尽管博伊克特的生活很简单,但他并不刻板。他对世界大事、科学界的社会政治抱有健康的兴趣,也喜爱美食和美酒。他总是第一个建议去酒吧。他对科学秉持着高标准,能做出激烈的批评,但他从不拒绝和任何人交流,无论对方是清洁工、博士后还是资历最高的教授,也从不挑话题,无论是闲聊、工作、政治还是深入的理论。如果你有想说的,他就会尊重地聆听。如果他之后觉得你的标准够不上他的,他还是会很友好,但他不会再多回应你的观点。他会避开大型科学会议,嘲弄人多的会议为“羊群”。他的讲座不会演练很久,通常很随意,有时很飘忽——不似现在这个作图容易的时代里那些华丽流畅、TED演讲风格的演示。
布赖恩会去接触任何他感兴趣的年轻科学家。他来美国时,经常会住到我在波士顿附近的小房子里。我们会在后廊上坐到晚上,谈论视网膜细胞的分类,聊一聊我们的朋友,喝一瓶波旁威士忌。布赖恩过世后,视网膜科学家团体投票为他的科学成就设立了一个纪念奖,两年一度在佛蒙特州的山上聚会并颁奖。因为布赖恩受到的尊敬和爱戴,博伊克特奖是一项为人珍视的荣誉;获奖者会被授予一张证书,以及一瓶单麦芽苏格兰威士忌。
[1] T.S.艾略特(T.S.Eliot)在诗作《荒原》的手稿中,借传说中的盲人先知忒瑞西阿斯之口写道:“在她脑中,可能飘过一个半成形的想法。”他的朋友庞德并不喜欢“可能”,并在页边空白处写下了这句尖刻的评论。对科学家和诗人来说,这都是个好建议——几乎知道是没有意义的。庞德的评论摘自《荒原》的这个版本:The Waste Land:A Facsimile and Transcript of the Original Drafts,ed.V.Eliot [New York:Houghton Mifflin,1971]
[2] 大多数细胞在自然情况下不会发光,科学家为了便于观察,会对动物或某些细胞做转基因操作,或者用下文所述的免疫组化方法直接染色,让它们在显微镜的激发光照射下发出荧光(fluorescence)。——译者注
[3] 通过追踪树突和轴突延伸的终点,你可以算出无长突细胞在视网膜中层细胞的占比,从而估算出无长突细胞的总量。在用分子标记染色的相片上,也可以用同样的方法估计每种已知类型的无长突细胞数量,加和之后,就可以知道已知的无长突细胞一共有多少,以及其占所有无长突细胞数量的比例。——译者注
[4] 恩里卡的原初想法是这样的:也许环境丰容有益于视觉,就像它有益于各种小鼠智力一样。她一发现确实如此,就做了更多的实验,来分离研究中的各个变量。Barone,I.,Novelli,E.,& Strettoi,E.(2014).Long term preservation of cone photoreceptors and visual acuity in rd10 mutant mice exposed to continuous environmental enrichment.Molecular Vision,20,1545–15
[5] 埃利奥·拉维奥拉可以称得上是高尔基染色的大师,因为他是在帕多瓦大学解剖学系接受的训练,而该系在19世纪就是卡米洛·高尔基之所在。
[6] 这个结论发表后的十年内,人们又用连续切片电子显微成像将瓦塞尔目录中的两种细胞进一步分成小类,所以一共有14种细胞。这部分只是个语义游戏,所以瓦塞尔的基本结论是正确的。Wässle,H.,Puller,C.,Müller,F.,& Haverkamp,S.(2009).Cone contacts,mosaics,and territories of bipolar cells in the mouse retina.Journal of Neuroscience,29,106–117.Helmstaedter,M.,Briggman,K.L.,Turaga,S.C.,Jain,V.,Seung,H.S.,&Denk,W.(2013).Connectomic reconstruction of the inner plexiform layer in the mouse retina.Nature,500,168–174.
[7] 这一节的人物侧写来自作者的个人接触,以及两本精彩的传记:Boycott,B.B.(2001).Brian B.Boycott.In Squire,L.R.,(Ed.).The history of neuroscience in autobiography.San Francisco:Academic Press.Wässle,H.(2002).Brian Blundell Boycott,10 December 1924–22 April 2000.Biographical Memoirs of Fellows of the Royal Society,48,51–68.










